文:回溯档案
编辑:回溯档案
了解开花和成熟的分子机制对于了解种子作物的物候和在给定环境下最大化产量非常重要,在拟南芥积累的知识基础上,人们对许多作物的开花分子机制进行了研究。

这些研究揭示了常见的重要基因,例如FLOWERING LOCUS T ( FT ) 和CONSTANS ( CO ),但也揭示了它们的功能差异和遗传机制的多样性,这些基因机制是物种内开花时间自然变异的基础 。

大豆是一种兼性短日照植物,光周期反应的丰富遗传变异使作物能够适应广泛的纬度,这种广泛的适应性是由控制开花的许多主要基因和数量性状基因座 (QTL) 的自然变异造成的。

迄今为止,已鉴定出 10 个控制大豆开花和成熟时间的主要基因而其他基因座的显性等位基因延迟开花和成熟,E6和J已在具有长幼熟习性的标准品种和晚花品种杂交的后代中被发现,这会导致短日照下的晚花 。

E9已通过对从野生大豆种质引入的早期开花 QTL 进行分子解剖来鉴定。涉及十个基因中的四个的分子机制已被确定。E1编码一种可能下调FT2a和FT5a的转录因子,并且对开花时间的影响最为显著。
E2是拟南芥 GIGANTEA ( GI )的直系同源物。E3E4和E4分别编码光敏色素 A 同工型 GmPHYA3 和 GmPHYA2 。
大豆基因组至少有 10 个FT同源物,其中 6 个在异位表达时促进拟南芥 ft突变体或生态型 Columbia的开花,它们的表达谱根据组织和生长阶段的不同而不同,表明它们在大豆开花中的亚功能化。

在六种同源物中,FT2a和FT5a已被广泛研究,因为它们的表达模式密切遵循光周期变化 并且它们的过度表达甚至在非诱导条件下也能促进开花。

FT2a和FT5a的光周期表达模式很可能受E1及其同源物E1La和E1Lb控制,而 E1La 和 E1Lb 又受E3和E4的控制。E2可能通过不同于 E1-PHYA 途径的途径抑制FT2a表达 。
E1 – E4的等位基因变异会导致大豆品种之间开花时间的部分差异,但不是全部差异,在E1、E3和E4处独立发生的各种突变组合会导致开花对光周期不敏感或低敏感度。
除了上述四个基因外,拟南芥开花基因的许多大豆直系同源物已被表征:COL、CRY , FKF1, FLD , FUL, RAV -类似(与 ABI3/VP1 相关-类似) , SOC1 / AGL20 ( COL1/AGAMOUS-LIKE 20 ) 、EAT1 靶标( TOE ) 和ZTL 过度表达的抑制剂。
一项全基因组关联研究还揭示了许多与开花时间显着相关的 SNP;其中一些 SNP 暗示拟南芥开花基因的直系同源物,参与控制大豆的开花时间。

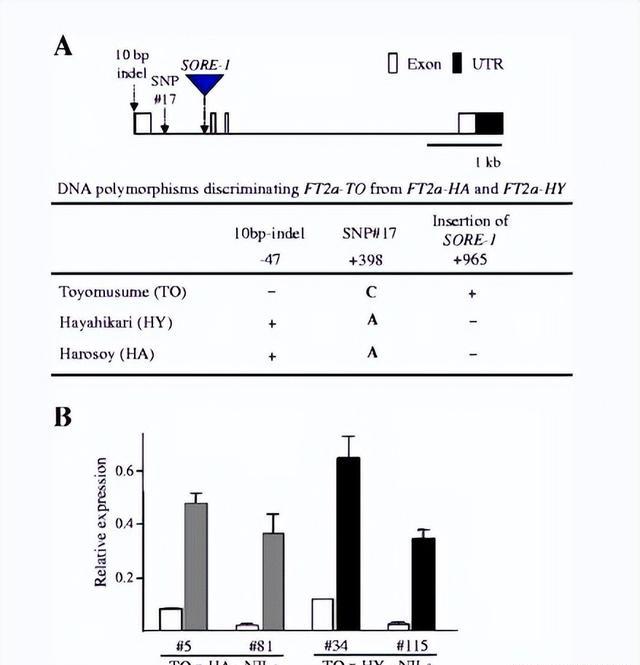
然而我们对这些直系同源物在大豆开花自然变异中的作用的理解仍然有限,在FT2a中发现了多种序列变异尽管编码区高度保守,但大豆品种中的启动子区仍然存在。
尽管其中一些多态性与所测试品种的开花时间变化显着相关,但它们在FT2a表达中的作用尚未完全了解 。
在这项研究中,利用不同来源的早熟品种之间的杂交,我们发现开花时间的分离部分与成熟基因 E9 的标签标记相关,我们证明E9与FT2a相同,其隐性等位基因在第一个内含子中插入了Ty1/copia样逆转录转座子,这降低了FT2a转录水平并延迟开花。
Harosoy 和 Toyomusume 杂交后代开花时间的分离
杂交中使用了两个早熟品种,加拿大品种和日本品种 。它们在E2、E3和E4处具有相同的成熟基因型,但E1基因型不同:HA 具有亚等位基因e1-as等位基因,而 TO 具有e1-nl等位基因,缺少包含整个E1基因的基因组区域 。

在日本札幌自然日照条件下,TO和HA几乎同时开花,但前者比后者早3至5天开花。然而,F 2群体的开花时间差异很大,由于E1等位基因变异对开花时间有很大影响,我们首先评估了E1等位基因对群体开花时间的影响。

我们使用等位基因特异性 DNA 标记 和侧翼简单序列重复 (SSR) 标记确定了F 2植物的E1基因型,正如预期的那样, e1-nl纯合植物(来自 TO)平均比e1-as纯合子。

由于每个等位基因纯合的植物在开花时间上仍然存在很大差异,因此我们对每个等位基因纯合的 16 株植物进行了后代测试。F 2个体的开花时间与其后代的平均开花时间密切相关。

e1-nl纯合子的亲代相关系数为 0.676 , e1-as纯合子的亲子相关系数为 0.823 ,这表明除E1之外的遗传因素在两个基因型类别中均存在分离。
测试开花时间和 SSR 标记之间的关联
为了检测独立于E1分离的开花基因,我们测试了每个e1-nl和e1-as基因型类别中的开花时间-SSR 标记关联;我们使用了位于拟南芥开花基因直系同源聚集的基因组区域的 61 个 SSR 标记。
两个标记与e1-nl纯合子中的开花时间显着相关,五个标记与e1-as纯合子中的开花时间显着相关,所有位点上 TO 等位基因纯合的植物 (A) 开花时间均晚于 HA 等位基因纯合的植物 。
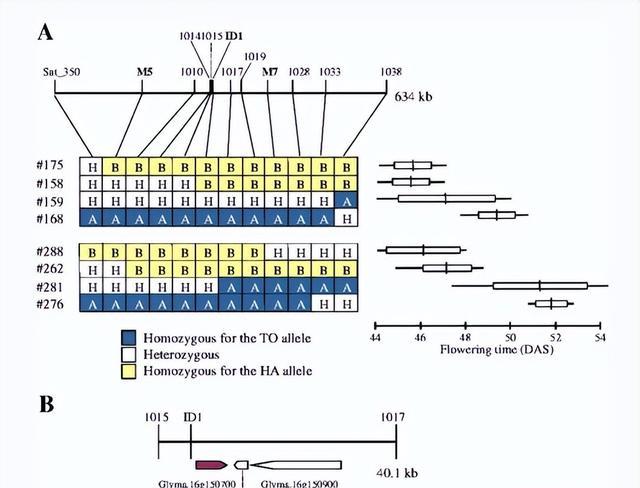
Sat_350 在e1-nl和e1-作为基因型类别,Sat_350 位于 LG J 上的 SSR 标记 Satt686 附近,该标记是在栽培大豆 (TK780) 和野生大豆 (Hidaka 4) 杂交中鉴定的E9基因的标签标记 的亲本。

而 TK780 携带隐性e9等位基因 ,因此 Sat_350 标记的基因可能与E9相同,并且 TO 与 TK780 具有相同的晚花隐性等位基因。
为了确认开花时间和ID1基因型之间的共分离,我们检查了14个e1-nl纯合的F 2家族和14个e1-as纯合的F 2 家族,在e1-nl家族中,TO等位基因纯合的两个家族的植物开花较晚,而HA等位基因纯合的两个家族的植物开花较早。

在 10 个杂合家族中观察到开花时间和标记基因型之间存在高度显着的关联同样,在具有e1-as基因型的 5 个杂合家族中,在开花时间和标记基因型之间检测到高度显着的关联,因此家族主要由 ID1 标记的基因型来解释。

FT2a基因组区域的序列分析


近等基因系和光周期不敏感种质中不同FT2 a 等位基因的表达
我们为来自 F 5杂合植物后代的上述三个FT2a等位基因开发了四组 NIL:两组来自 TO 和 HA 之间的杂交,两组来自 TO 和 HY 之间的杂交。

我们发现,在SD条件下,FT2a - TO的表达量远低于FT2a - HA和FT2a - HY的表达量。
接下来,我们检查了NIL 杂合兄弟姐妹中FT2a 的表达;该分析基于以下事实,FT2a-HA中创建了Dde I 限制性位点,但在FT2a-TO和FT2a-HY中则没有。
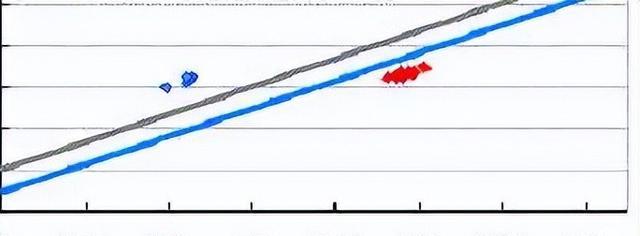

杂合子中略低,纯合子中非常低,PCR产物消化显示,在杂合子中, FT2a-HA的转录水平远高于FT2a-TO,这种差异表明FT2a-TO的较低表达是由等位基因特异性转录抑制引起的。
而不是由 RNA 沉默的序列特异性 RNA 降解引起的,RNA 沉默降低了两个等位基因的转录物水平。
成熟基因E9是FT2a
TO 和 HA 与E1和E9基因座等位基因共分离的杂交F 2和 F 3后代的开花时间,精细定位将E9界定为包含三个基因的40.1-kb区域,其中包括FT2a 。

FT的大豆直系同源物。测序和表达分析表明FT2a最有可能是E9的候选者,并且e9导致的延迟开花很可能是由FT2a减少引起的,转录丰度。


TO 与 HA 和 HY 的区别还在于第一个内含子中的SNP 和SORE-1插入,其中后者仅与FT2a表达水平相关,因此我们的研究表明,SORE-1的插入减弱了FT2a 的表达并延迟了开花。

大豆基因组共有10个FT直系同源物,其中6个保留了FT功能,可以促进拟南芥ft突变体或Col-0开花,因此所有六种同源物都可以作为大豆中潜在的花诱导剂。
尽管只有其中两种,FT2a和FT5a,在开花分子机制的研究中得到了广泛的表征这项研究表明,不同水平的FT2a表达直接调节大豆开花时间的自然变化。

导致FT2a表达减弱的因素
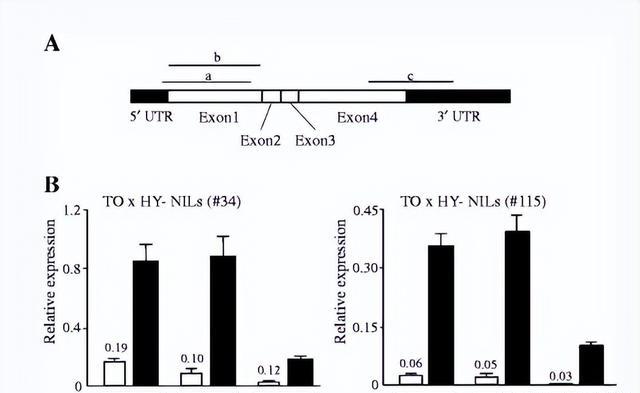
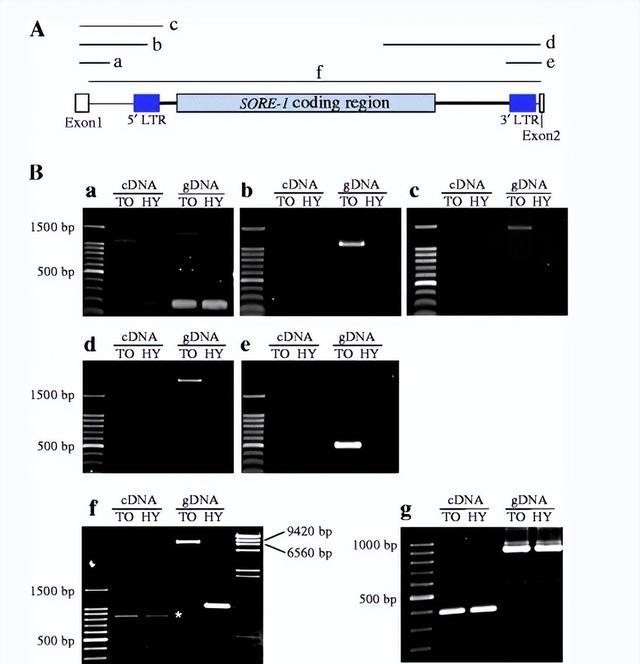
插入内含子的植物 TE 可能会影响 RNA 加工并使其宿主基因容易受到短干扰 RNA (siRNA) 介导的沉默影响,我们的结果显示第一个内含子(包括SORE-1)被剪接掉。

因为在检测到剪接产物时没有检测到 覆盖FT2a外显子和SORE-1的初级RNA转录本,因此SORE-1插入不会显着干扰FT2a RNA 加工。
结论
本研究表明,大豆成熟基因E9是拟南芥 FT的直系同源基因FT2a,其隐性等位基因通过较低的转录本丰度延迟开花,因此 FT2a直接参与大豆开花时间的自然变化。
FT2a表达的减弱是由第一个内含子中插入SORE-1引起的等位基因特异性转录抑制引起的,隐性e9等位基因是泄漏等位基因;其由参与光周期响应的其他基因的调节被保留。
因此,它可以在早花遗传背景下维持营养生长,并且也可作为低纬度地区品种发育中的长幼年等位基因,这些地区强烈促进开花。
参开文献
【1】Andrés F,Coupland G。开花对季节线索反应的遗传基础。纳特·吉内特 (Nat Rev Genet)。2012;13:627–39。
【2】Itoh H,Izawa T。水稻长日照条件下成花素基因表达和花转变的临界日长识别的一致性。摩尔工厂。2013;6:635–49。
【3】Pin PA,Nilsson O。FLOWERING LOCUS T在植物发育中的多方面作用。植物细胞环境。2012;35:1742–55。
【4】Watanabe S、Harada K、Abe J。大豆开花光周期响应的遗传和分子基础。品种科学。2012;61:531–43。
【5】伯纳德·RL. 大豆开花时间的两个基因。作物科学。1971;11:242–4。
