文:回溯档案
编辑:回溯档案
棉花纤维是从受精胚珠延伸出来的种子毛,棉纤维是最长的单细胞之一,可以长到6厘米,植物激素直接影响棉纤维细胞的生长和延伸,众所周知,生长素和赤霉素可促进纤维细胞的形成和发育。

对未成熟胚珠和含纤维胚珠表达序列标记(ESTS)的测序分析揭示了与 自适应因子 ( 阿尔夫 )及赤霉素信号,葡萄糖类固醇和乙烯也能积极影响纤维的发育,而脱落酸和细胞分裂素抑制纤维细胞发育。

此外编码假定的myb转录因子的棉花基因在纤维发育的早期阶段就会被诱导,但在一个在纤维形成过程中受到损害的裸种子突变体中受到抑制。
这些数据与已知的肌酸和其他转录因子在叶毛发育中的作用相符及棉纤维发,许多编码假定转录和植物激素反应因子的基因是微转录核的目标。

小型干扰性的基因表达和表观调控是控制基因表达和表观遗传调控的多样物种中产生的21-24核苷酸小型的基因表达调控,另外植物生产 特朗斯 -代理家长,由压力引起的自然反义情绪,及病原体诱导的长年西纳。
由RNA聚合酶II转录成主要的米尔纳转录酶,这些转录酶由类似核细胞的酶处理,如动物体内的迪克尔酶和罗沙酶处理以及植物中的类似双星蛋白。

成熟的微核反应堆通常通过计算分析和/或实验方法(如克隆和测序)来识别,截至2009年3月v,米尔贝基数据库发布13.0,包含3,788个植物MIRNA条目。
虽然许多转录因子和植物激素因子是棉棉纤维发育的目标,并预计将在棉棉纤维的发育中发挥作用,但棉花中的小RNA数据有限,部分原因是棉花基因组序列不存在。

经对棉花无害环境技术进行计算分析后,只确定了十几个多边环境影响评估及低排量测序,较少的先质结构沉积在米尔基,最近一项利用高吞吐量测序的研究发现,棉花中有34个保守的微核系统和8个最低编码的微核系统。
为了丰富棉花纤维发育中的小RNAS知识,我们分析了纤维和胚珠发育早期的MRNAS,我们对棉叶、未成熟胚珠和含纤维胚珠中大约400万个小胚珠进行了排序和分析。

棉花中24核肽小胚珠在含纤维胚珠中含量较高,我们发现了27个受保护的家庭,确定了4个新的家庭,并预测了代表19个独一无二的家庭的32个米尔纳前体。
总共223个米尔纳目标被计算预测,其中一个子集被实验验证。在纤维发展的早期阶段,包括新的微纳米体在内的许多微纳米体都受到抑制,这与八个被测试目标的上浮一致,在纤维细胞的快速发育过程中,纤维带纤维胚珠中SRNAS的丰富和微生物的降低是小基因介导的重要因素。
棉花中的小RNAS分布情况
为了对棉花中的小胚珠进行表征,我们利用从叶子中提取的总RNA、未成熟胚珠(开花前3天,-3da)、带纤维细胞首字母的胚珠(花期当日,0da)和幼小含纤维胚珠(花期后3天,+3dd)建立了四个条码测序库。

在移除适配器序列后,我们确定了与已知细胞小RNAS、线粒体和质体序列相匹配的读取(约6%),大量的生读,从胚珠的6.4%到叶的53.8%不等,与Rrna相匹配。
这表明在树叶中降解的Rrna比例很高,或者,树叶中的rrna基因可能会通过rna介导的途径受到沉默或核仁支配。

棉花胚珠中最丰富的小核苷酸为24个长核苷酸。
棉花中最丰富的小型核苷酸是24个核苷酸,其次是26个核苷酸或更长的核苷酸和22个核苷酸,有意思的是,胚珠(0dA)和含纤维胚珠(+3dA)中,78%至84%的小核苷酸长为24个核苷酸长。
在拟南芥 ,24核肽小型RNAS在叶片中的分布约为43%,在花序中约为61%,在种子中约为41%。24个核苷酸的小RNAS主要包括与重复子和转座有关的SRNA。
高水平的24核苷酸小RNas 拟南芥 花序和发育的棉花胚珠与那些 拟南芥 而棉叶可能表明这些元素在胚珠或花序中受到抑制,或者,重复和其他元素通常在叶子中被抑制,但在细胞快速发育期间被激活。棉花基因组测序后,24核肽小RNAS对纤维发育的影响仍有待研究.

棉花中的微营养不良物质的识别
我们采用了共同标准以识别棉花中已知的微生物多样性和(或)前体,首先,棉质米尔纳必须与其他物种的异种异种异种亚种具有序列守恒和同源性,第二,如果米尔纳与已知的无害环境技术相匹配,则茎环结构清楚地显示米尔纳和米尔纳*在双工结构的另一端。

四倍体棉棉中细胞核核的茎环结构及新细胞核核的鉴定
发夹柱环结构是使用SIDR图表工具在未折叠包装中可视化的,利用米尔切克方法,识别了32个米尔纳前体,包括19个独特的家庭,代表的只是小部分的MIRNA家族在本研究中确认。

这些保守的结构特征已被建议用以指导由dcl-调解的米尔纳前体的加工. 格拉米尔156 发现有一些不同的特征,比如在MIRNA*中的一个鼓起的G,表明 Gumr156 匹配来自于 G.arboreum -与次基因组似 G.hirsutum 。


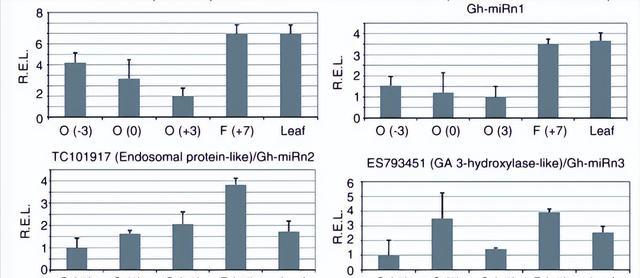
此外,MR2948有MR2948*,与MR29482密切匹配,表明MR2948可能是从MR29484家族进化而来的,预测GH-MR2948靶点中的一个将编码蔗糖合成酶样基因。

测序显示未成熟胚珠和含纤维胚珠的GH-MER2948水平均低于叶,这与纤维细胞发育早期阶段蔗糖合成酶基因(U73588)的生长有关.
棉花中保守微生物的差异表达
为进一步研究棉花微核的特性,我们采用了微RNA微阵列 测定棉纤维和非纤维组织中的微RNA积累模式,每台微阵列对85个不同的原子能机构和23个原子能机构进行了审讯以及26。

在棉花中,共有111个由这些物种27个科组成的毫雷纳夏在检测水平以上表示,微阵列结果证实了测序库中存在的21个保守的微磁核的表达,并在一个或多个组织中,显示另有55毫米,包括: G.hirsutum L.cv。
无纤维胚珠(o;+3d)的 N1N1 TM-1背景下纤维产量下降的裸种子突变体虽然基于跨物种杂交的检测可能会引入假阳性,但在微阵列中检测到的其他微纳米反应堆表明,本研究中确定的微纳米反应堆是不饱和的。

通过直接切取mrna来调控基因表达或抑制mrna翻。为测试棉质中的微纳米反应堆是否触发了目标分裂,我们使用Perl脚本和前面描述的目标预测标准预测了微纳米反应堆的目标。
米尔纳目标通常被发现在反意义的打击与三个或更少的不匹配的标准米尔纳序列之外,共为31个假定的米尔纳家庭确定了233个潜在目标,其中包括27个保护家庭和4个新的家庭。

许多预测目标均源自纤维发展早期阶段的无害环境技术,与…相比 拟南芥 ,棉花中每个MIRNA的预测靶点数量很大,这意味着在这个异体四倍体物种中有额外的类似和同源的基因。

纤维发育早期24核苷酸小RNAS的积累
高通量测序分析表明,在纤维和胚珠发育的早期阶段,24核苷酸的小RNAS爆发,虽然在这些组织中,由于微核细胞的数量相对较少,其他小的微核细胞的读数可能会增加,但微核细胞的总读数相对较小。

24核肽小rNas的数量占含纤维胚珠小RNA总读量的78%至84%,但只有大约50%的叶片和大约57%的未成熟胚珠,大多数24核苷酸小RNA都是从内源性重复基因衍生出来的,被称为重复相关的SRNA。

结论
对大规模并行测序数据、米尔纳微阵列、小RNA斑点和靶向分裂试验的分析表明,基因表达和生理学的快速动态变化与24核肽的普遍浓缩有关,并与异体四倍体棉花胚珠和纤维发育过程中的抑制有关。
小规模的棉花最低生活标准来自于大规模棉花最低生活标准集合的一小部分,但与相当大数量的世界最低生活标准追踪数据相匹配, G.raimondii 基因组。
在纤维发育过程中,胚珠和非纤维组织中的SRNAS的丰富表明,在纤维发育过程中,小RNA代谢和染色质修饰是活跃的,在纤维中,包括新的微核抑制在内的微核抑制与编码转录和植物激素反应因子的12个经验证的微核靶标的上位作用相关,包括以下基因: 阿尔夫 和 苏西3 在棉纤维中有高度表达。
这项工作提供了基因组和基因表达数据的丰富来源,并提供了一些新的发现,包括4个新的微核反应堆、一个新的候选人微核反应堆、25个新的微核反应堆前体和200多个预测目标,这将有助于今后研究微核反应堆和小核反应堆在棉花纤维发展中的作用。
参考文献
【1】糖合酶基因表达抑制抑制棉花纤维细胞的产生、延伸和种子发育。植物细胞。2003年,15:952-964。
【2】奥克尔曼MJ,SAKAIH:通过一个微子及其顶-2样靶基因调节开花时间和花器官特性。植物细胞。2003,15:273-2741。
【3】莫舍拉、麦尔尼克、凯利卡、邓恩雷姆、斯图霍姆DJ、鲍尔康贝(DC):单亲本的表达 拟南芥 .Nature.2009,460:283-286。
【4】莫尔纳、施瓦奇、斯图霍尔梅、苏灌曼、鲍尔康、DC:MIRNAS控制单细胞藻类基因表达 赖纳蒂 .Nature.2007,447:1126-1129。
【5】雷斯脱博士,罗斯jj,戴维斯pj,里德jb:孟德尔的茎长度基因编码赤霉素3β羟基化酶。植物细胞。1997年,9:1435-1443。
