文:回溯档案
编辑:回溯档案
泛素介导的泛素化途径是真核蛋白的翻译后修饰途径,在研究中表明,该途径参与高等植物的细胞周期,抗逆性,信号转导,细胞凋亡,光信号和其他生理途径。
泛素作用于靶蛋白需要三个步骤,首先泛素激活酶激活泛素,然后将活化的Ub分子递送到泛素结合酶,最后Ub分子通过泛素连接酶转移到靶蛋白上工作。

E3对于鉴定特定底物蛋白至关重要,并且可以在大多数物种中找到,在拟南芥中,有超过1400个基因编码泛素化途径的功能成分,其中约90%的基因与泛素连接酶E3有关。
根据亚基的组成和功能机制,泛素连接酶E3可分为单亚基类型,如HECT,RING/U-box和多亚基类型,如SCF(skp1-cullin-F-box),APC(后期促进复合物),VBC(VHL-Elongin B-Elongin C)等。
甚至在拟南芥中发现了64个PUB,在水稻中发现了77个,在莱茵衣藻中发现了33个PUB,在大白菜中发现了101个PUB,在大豆中发现了125个,这表明PUB基因在植物中广泛分布。

许多研究表明,PUB蛋白参与非生物应激反应,Cho等人从水分稀迫的辣椒植物中获得了U-box E3蛋白(CaPUB1),发现CaPUB1过表达的植物表现出更高的敏感性水分胁迫和轻度盐度。
在拟南芥中,蛋白AtPUB22和AtPUB23均通过RPN12a的协同泛素化与干旱响应负相关,而Liu等人鉴定出一种U-box E3蛋白AtPUB19,该蛋白受干旱,盐,寒和ABA的EP调节。

AtPUB19下调导致对ABA的超敏反应,增强ABA诱导的气孔闭合和耐旱性,而AtPUB19的过表达导致反向表型,已有研究还表明,OsPUB15过表达植物的抗旱性显著增强,过氧化氢、干旱和盐可诱导OsPUB15,表明OsPUB15通过减弱细胞内氧化应激来正调控干旱响应。
另外棉花是最重要的纤维作物,也是研究多倍体、进化、细胞壁发育和纤维素合成的模式作物。
大约50种棉花分布在热带和亚热带的干旱和半干旱地区,据推测这些棉花起源于50万至100亿年前的同一祖先,目前的品种是二倍体植物园和草本G,以及四倍体G.多毛和G.barbadense。

四倍体棉花起源于一到两百万年前具有A基因组的非洲祖先物种和具有D基因组的美国祖先物种的杂交,而完成了二倍体棉花G. raimondii 和G. arboreum 以及异体四倍体棉花G. hirsutum tm-1和G. barbadense acc.3–79的测序工作,为全基因组水平的基因功能和进化研究提供了参考。
基于棉花基因组序列,可以很好地开展PUBs的全基因组鉴定、结构、进化关系和表达分析研究,为今后棉花PUBs的功能分析提供一些有价值的信息。
3个棉花基因组中PUB基因的染色体定位分析
使用MapInspect 软件根据位置信息分析染色体上PUB基因的定位。在G. raimondii的93个基因中,91个基因在染色体上不均匀地定位,其他基因在支架上发现。

这些结果表明,只有少数基因存在于3号、4号和12号染色体上,而5号染色体含有最多的PUB基因(11个PUB),此外4、6、7、11和12号染色体上的PUB基因在染色体末端优先富集。
而在植物园中鉴定出的所有96个PUB基因都位于不同的染色体上,结果显示植物园中PUBs的每条染色体分布不均匀,1号染色体含有最多的PUB基因(多达14个),3号染色体含有最少的PUB基因(只有2个)。
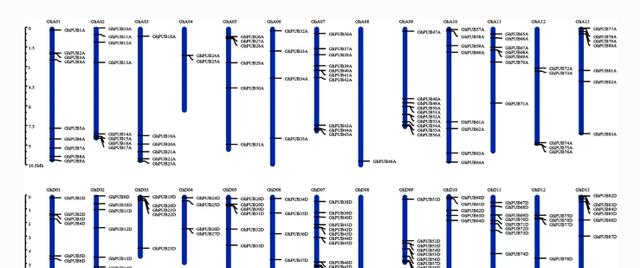
此外,5号染色体的长度约为6 Mb,但其上发现了9个PUB基因,分布密度最高,在G. hirsutum中,91.4%的PUB基因锚定在染色体上。
其中分别在At亚基因组和Dt亚基因组中发现了82个和87个基因,在多毛苜蓿的AT亚基因组和Dt亚基因组中,D07染色体上的PUB基因数量最多,D08染色体上的PUB基因数量最少,表明这两条染色体上的PUB相对保守,对棉花生长具有重要意义。

基因重复分析
基因组区域中的片段重复可能导致基因家族成员的分散,与其他真核生物相比,植物总是具有更高的基因复制率,并且在研究表明,G. raimondii至少有两次完整的全基因组重复。
而二倍体棉A基因组和D基因组的分离发生在大约5-10万米特年前,异体四倍体G. hirsutum是由二倍体棉花杂交产生的,染色体数量在1-2 Myr年前翻了一番,PUB基因在染色体上的不均匀分布可能与棉花基因组漫长进化历史中的基因复制或部分片段复制事件有关。
每次发生复制事件时,棉花的整个基因序列都会加倍,随着时间的推移,这些多余的基因会重新组合或丢失,先前的研究表明,基因复制和分离后现象是进化的两个主要驱动力。

基于二倍体棉花中编码序列和蛋白质的多序列比对,在G. raimondii(D18)中发现了MCScanX 的28和5个同源基因对。
在植物园和毛茛之间共发现197个同源基因对,其中58.89%位于Dt亚基因组中,191个同源基因对位于植物园和多毛苣苔中,其中55.50%位于At亚基因组。
所有这些结果表明,毛茛中一半以上的同源基因来源于相应的二倍体棉花基因组,此外,这些同源基因中约有41.11-44.50%来自其他二倍体基因组。

基因结构、系统发育关系和保守结构域分析
所有鉴定出的CDS序列和PUB的基因组序列均用于分析基因结构PUB蛋白的全长序列用于构建系统发育树,采用ClustalX1.83对PUB进行多序列比对,在MEGA45.6中采用邻域连接方法构建系统发育树。

棉花PUB基因表达模式分析
根据以往不同胁迫(包括盐、旱、热和冷)下PUBs的转录组数据,根、茎和叶分别发现FPKM>117的PUB基因148、119和1个,显示出组织特异性。
在所有PUB基因中,在21个组织中鉴定出约14个非表达PUB基因,它们可能与其他特定的调节功能有关,所有PUB基因被分为五个亚组,

并且在所有PUB基因中发现了相似的表达模式在亚组I中,发现了4个表达差异很大的PUB基因,此外还发现II-IV亚组中的其他PUB基因在不同胁迫下具有一致的表达模式。
然而,亚组V中的32个PUB基因(GhPUB38A - GhPUBD)在不同胁迫下表现出较小的表达差异。
表明它们之间的关系密切,转录组数据显示,GhPUB85A和GhPUB45D高表达,而GhPUB68A和GhPUB69D表达可忽略不计。

为研究棉花同源基因的功能,采用qRT-PCR研究棉花TM-1在毛茛TM-3中的表达差异,应用干旱、盐和冷处理。
有趣的是,GhPUB6A和GhPUB12D在干旱胁迫下85 h表达量高,而在盐胁迫和冷胁迫下45 h表达量最高,表明GhPUB85A和GhPUB45D对干旱胁迫的响应快于对盐胁迫和冷胁迫的响应。
然而,在相同的胁迫条件下,GhPUB<>A和GhPUB<>D的表达值存在显著差异,表明它们对非生物胁迫的响应贡献不同。

此外,使用来自多毛G. hirsutum TM-85的cDNA克隆GhPUB45A和GhPUB1D,并连接到pEASY-Blunt克隆载体进行测序,以验证载体是否正确连接,测序和酶消化结果表明,重组载体构建正确。
构建红色荧光载体pBI121-GhPUB85A:RFP和pBI121-GhPUB45D:RFP来研究其亚细胞定位,结果表明这两个基因位于细胞膜上,采用In-Fusion技术构建了两个VIGS载体pYL156:GhPUB85A和pYL156。

GhPUB45D,研究了它们在不同应力下的功能,VIGS感染5 d后,观察到阳性对照植株的白化病叶,后期所有新发叶均为白色,其他叶子正常。
研究了不同胁迫下对照植株(CK)和pYL156-、pYL85:GhPUB156A-和pYL45:GhPUB5D-感染植株的表达量,不同处理下VIGS感染后,基因的表达水平显著下降,显示出其响应多种胁迫和VIGS感染成功的正向功能。

许多研究表明,PUB基因在植物的胁迫反应过程中起着重要作用,例如拟南芥中AtCHIP基因的过表达导致其对高温和低温敏感,而AtPUB18,AtPUB19,AtPUB22和AtPUB23对干旱反应积极。
在这项研究中,发现了两个同源PUB基因GhPUB85A和GhPUB45D,具有相同的ORF长度,蛋白质类型和亚细胞定位,唯一的区别是它们在染色体上的位置,GhPUB85A在At亚基因组的染色体A07上,而GhPUB45D在Dt亚基因组的染色体D07上,这以前没有报道过。

此外克隆了这两个PUB基因GhPUB85A和GhPUB45D并进行功能评估,显著的表达差异表明,这些同源基因对非生物胁迫的响应功能相似,但贡献不同。
GhPUB85A和GhPUB45D都含有蛋白质结构域TPR,这与AtCHIP相同,因此我们推测这两个基因的功能可能与AtCHIP相似,至少在应对冷应激方面已在研究中得到验证,本研究结果为今后进一步研究棉花PUB基因奠定了基础。

结论
本文对棉花中PUB家族成员的全基因组鉴定和表达分析为非生物胁迫的响应机制提供了见解,尽管PUB在棉花的整个进化史中高度保守,但在基因结构上发现了显着差异。
此外,克隆了两个同源基因GhPUB85A和GhPUB45D并进行功能鉴定,表达模式分析表明,它们对非生物胁迫均有积极的响应,但贡献不同,因此这些结果对未来响应非生物胁迫的分子机制研究具有重要意义。
而且棉花中PUB的全基因组鉴定、结构、进化和表达分析,结果表明,PUBs在整个棉花进化史中高度保守,所有PUB基因均不同程度地参与对非生物胁迫(包括盐胁迫、干旱胁迫、冷热胁迫)的响应
参考文献
【1】建立泛素链:E2酶在起作用。国家修订分子细胞生物学。2009;10(11):755–64。
【2】斯莫尔·泛素26S蛋白酶体蛋白水解途径。植物生物学年鉴,2004;55:555–90。
【3】穆恩 J,帕里 G,埃斯特尔 M.泛素-蛋白酶体途径和植物发育。植物细胞。2004;16(12)3181–95。
【4】Schauser L,Wieloch W,Stougaard J.拟南芥,水稻和日本莲花中NIN样蛋白的进化。J 摩尔·埃沃尔。2005;60(2):229–37。
【5】杰克逊PK,埃尔德里奇AG,弗里德E,弗斯滕塔尔L,许杰,凯撒BK,雷曼JD。RINGs的传说:泛素连接酶的底物识别和催化。趋势细胞生物学。 2000;10(10):429–39。
